|
Cell death extends beyond loss of viability, involving oxidative stress, membrane injury, metabolic shifts, and ATP release. Combining multiple readouts better captures how dying cells affect their environment. In a cancer model, immunogenic death of 4T1 cells increased ROS and ATP release, released nuclear damage-associated proteins, and exposed immunogenic surface markers, promoting dendritic cell maturation and CD8 T cell activation. In a neural model, Friedreich's ataxia microglia induced neuronal Fe²⁺ accumulation, oxidative DNA damage, and elevated apoptotic markers. These findings reflect cell death as a stress-driven process that reshapes its surrounding environment.
|
 |
Programmable nanomicelles rewire myeloid immunity for durable control of primary and metastatic breast cancer
(Yang et al., Nature Communications, 2026)
Summary:
This study shows that cell death involves more than a drop in viability. In 4T1 breast cancer cells undergoing immunogenic cell death, ROS and extracellular ATP release increased, HMGB1 was released as a tumor damage signal, and calreticulin moved to the cell surface as a cue for antigen-presenting cells. Conditioned media from these cells promoted dendritic cell maturation and enhanced the tumor-attacking activity of CD8 T cells.
Highlighted technique:
To distinguish simple cytotoxicity from immune-relevant cell death, the authors focused on 4T1 cell viability, intracellular ROS, and extracellular ATP release. These were measured using a CCK-8 assay, DCFH-DA fluorescence imaging, and ATP release quantification, providing readouts for cell death extent, oxidative stress, and ATP released from damaged tumor cells.
Cell viability helps quantify the extent of cell death, while indicators such as intracellular ROS and extracellular ATP provide additional readouts of stress-associated signals accompanying cell death.
Microgliopathy as a primary mediator of neuronal death in models of Friedreich’s Ataxia
(Pernaci et al., Nature Communications, 2025)
Summary:
This study examined whether microglial abnormalities seen in Friedreich's ataxia (FRDA) are merely a consequence of neuronal death or can act as a causal driver. Disease-derived microglia showed mitochondrial impairment, increased ROS, iron accumulation, lipid peroxidation, and reduced lysosomal function. In 2D co-culture with healthy neurons, this led to neuronal iron accumulation, oxidative DNA damage, elevated markers of apoptotic execution, and associated morphological changes. These findings indicate that microglial dysfunction can directly drive neuronal injury, rather than merely arising as a downstream consequence of it.
Highlighted technique:
To evaluate intercellular stress preceding neuronal injury, the study assessed mitochondrial ROS in FRDA iPSC-derived microglia by fluorescence imaging. In 2D co-culture with healthy neurons, it measured neuronal Fe²⁺ accumulation and evaluated DNA damage, including γH2AX and other oxidative damage markers.
To evaluate intercellular stress that precedes neuronal injury, it is useful to look beyond cell viability alone and assess microglial mitochondrial ROS together with neuronal intracellular Fe²⁺ levels and γH2AX-based DNA damage.
Cell Death Related Activity Indicators (click to open/close)
Application Note I (click to open/close)
> Evaluation using Staurosporine-treated cells
HeLa and A549 cells were treated with Staurosporine, and extracellular ATP release and cell viability were assessed over time. The results showed that the extracellular ATP release of each cell differed in amount and timing depending on the cell type.
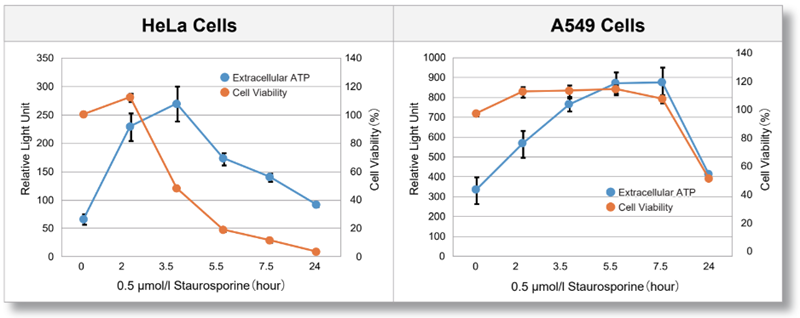
<Product in use>
Extracelluar ATP : Extracellular ATP Assay Kit-Luminescence (code:E299)
Cell viability : Cell Counting Kit-8 (code:CK04)
Application Note II (click to open/close)
> Accurate Autophagy Imaging with Lysosome Staining
|
|
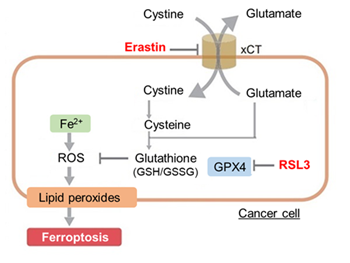
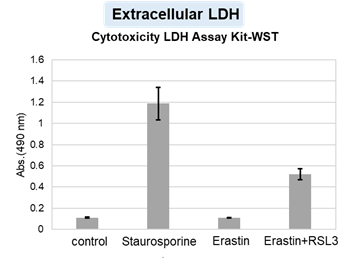
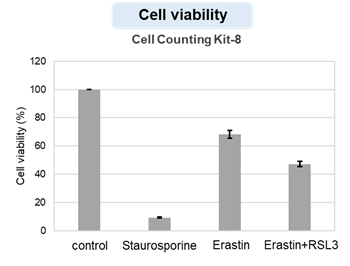
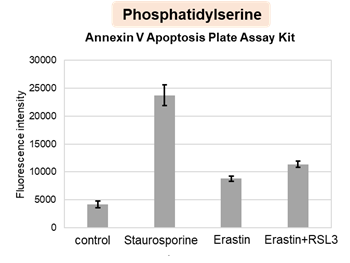
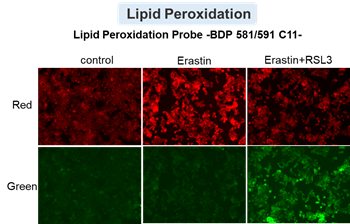
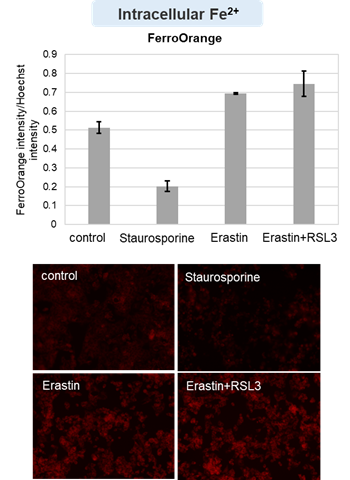
HepG2 cells treated with the apoptosis-inducing agent staurosporine or the ferroptosis-inducing agents Erastin and RSL3. After treatment, extracellular LDH, phosphatidylserine, cell viability, intracellular Fe2+ and lipid peroxidation were determined.
The results showed that apoptosis-induced cells treated with staurosporine showed an increase in phosphatidylserine, a decrease in cell viability and an increase in extracellular LDH, indicating that cell death had occurred. On the other hand, intracellular Fe2+, an indicator of ferroptosis, remained unchanged. In cells treated with Erastin, a ferroptosis inducer, intracellular Fe2+ increased and cell viability decreased, but extracellular LDH and lipid peroxidation did not increase. In cells in which ferroptosis was more strongly induced by co-treatment with RSL3 in addition to Erastin, increased intracellular Fe2+ and lipid peroxidation were observed. Moreover, decreased cell viability and increased dead cells were detected. Meanwhile, phosphatidylserine showed a lower rate of increase during ferroptosis induction compared to apoptosis-induced cells. These results suggest that cell death can be distinguished by evaluating a combination of cell death indicators.
|
 |
|
 |
|













